pits on the radial walls. The large medullary rays give to the wood
a characteristic parenchymatous or lax appearance, which is in
marked contrast to the more compact wood of a conifer. The
protoxylem-elements are situated at the extreme inner edge of the
secondary wood, and may occur as small groups of narrow, spirally-pitted
elements scattered among the parenchyma which abuts on the
main mass of wood. Short and reticulately-pitted tracheal cells,
similar to tracheids, often occur in the circummedullary region of
cycadean stems. In an old stem of Cycas, Encephalartos or Macrozamia
the secondary wood consists of
several rather unevenly concentric
zones, while in some other genera it
forms a continuous mass as in conifers
and normal dicotyledons. These
concentric rings of secondary xylem
and phloem (fig. 9) afford a characteristic
cycadean feature. After the
cambium has been active for some
time producing secondary xylem and
phloem, the latter consisting of sieve-tubes,
phloem-parenchyma and frequently
thick-walled fibres, a second
cambium is developed in the pericycle;
this produces a second vascular
zone, which is in turn followed by a
third cambium, and so on, until several
hollow cylinders are developed. It
has been recently shown that several
cambium-zones may remain in a state
of activity, so that the formation of a
new cambium does not necessarily
mark a cessation of growth in the
more internal meristematic rings. It
occasionally happens that groups of
xylem and phloem are developed
internally to some of the vascular
rings; these are characterized by an
inverse orientation of the tissues,
the xylem being centrifugal and the
phloem centripetal in its development.
The broad cortical region, which contains
many secretory canals, is traversed
by numerous vascular bundles (fig. 9, c) some of which pursue
a more or less vertical course, and by frequent anastomoses with one
another form a loose reticulum of vascular strands; others are leaf-traces
on their way from the stele of the stem to the leaves. Most of
these cortical bundles are collateral in structure, but in some the xylem
and phloem are concentrically arranged; the secondary origin of
these bundles from procambium-strands was described by Mettenius
in his classical paper of 1860. During the increase in thickness of a
cycadean stem successive layers of cork-tissue are formed by phellogens
in the persistent bases of leaves (fig. 9, pd), which increase in size
to adapt themselves to the growth of the vascular zones. The leaf-traces
of cycads are remarkable both on account of their course and
their anatomy. In a transverse section of a stem (fig. 9) one sees
some vascular bundles following a horizontal or slightly oblique
course in the cortex, stretching
for a longer or shorter
distance in a direction concentric
with the woody
cylinder. From each leaf-base
two main bundles
spread right and left
through the cortex of the
stem (fig. 9, lt), and as they
curve gradually towards the
vascular ring they present
the appearance of two
rather flat ogee curves,
usually spoken of as the
leaf-trace girdles (fig. 9, lt).
The distal ends of these
girdles give off several
branches, which traverse
the petiole and rachis as
numerous collateral bundles. The complicated girdle-like course is
characteristic of the leaf-traces of most recent cycads, but in some
cases, e.g. in Zamia floridana, the traces are described by Wieland
in his recent monograph on American fossil cycads (Carnegie Institution
Publications, 1906) as possessing a more direct course similar to
that in Mesozoic genera. A leaf-trace, as it passes through the cortex,
has a collateral structure, the protoxylem being situated at the inner
edge of the xylem; when it reaches the leaf-base the position of the
spiral tracheids is gradually altered, and the endarch arrangement
(protoxylem internal) gives place to a mesarch structure (protoxylem
more or less central and not on the edge of the xylem strand). In a
bundle examined in the basal portion of a leaf the bulk of the xylem
is found to be centrifugal in position, but internally to the protoxylem
there is a group of centripetal tracheids; higher up in the petiole the
xylem is mainly centripetal, the centrifugal wood being represented
by a small arc of tracheids external to the protoxylem and separated
from it by a few parenchymatous elements. Finally, in the pinnae of
the frond the centrifugal xylem may disappear, the protoxylem being
now exarch in position and abutting on the phloem. Similarly in
the sporophylls of some cycads the bundles are endarch near the base
and mesarch near the distal end of the stamen or carpel. The
vascular system of cycadean seedlings presents some features worthy
of note; centripetal xylem occurs in the cotyledonary bundles
associated with transfusion-tracheids. The bundles from the
cotyledons pursue a direct course to the stele of the main axis, and
do not assume the girdle-form characteristic
of the adult plant. This
is of interest from the point of view
of the comparison of recent cycads
with extinct species (Bennettites), in
which the leaf-traces follow a much
more direct course than in modern
cycads. The mesarch structure of
the leaf-bundles is met with in a less
pronounced form in the flower peduncles
of some cycads. This fact is
of importance as showing that the
type of vascular structure, which
characterized the stems of many
Palaeozoic genera, has not entirely
disappeared from the stems of modern cycads; but the mesarch bundle
is now confined to the leaves and peduncles. The roots of some cycads
Roots.
resemble the stems in producing several cambium-rings;
they possess 2 to 8 protoxylem-groups, and are
characterized by a broad pericyclic zone. A common phenomenon in
cycads is the production of roots which grow upwards (apogeotropic),
and appear as coralline branched structures above the level of the
ground; some of the cortical cells of these roots are hypertrophied,
and contain numerous filaments of blue-green Algae (Nostocaceae),
which live as endoparasites in the cell-cavities.
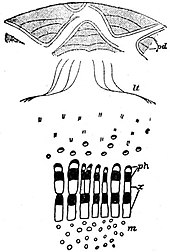
|
| Fig. 9.—Macrozamia.
Diagrammatic transverse
section of part of Stem.
(After Worsdell.)
pd, Periderm in leaf-bases.
lt, Leaf-traces in cortex.
ph, Phloem.
x, Xylem.
m, Medullary bundles.
c, Cortical bundles.
|
|
|

|
|
Fig. 11.—Ginkgo adiantoides.
Fossil (Eocene) leaf from the
Island of Mull.
|
|
|

|
| Fig. 10.—Ginkgo biloba. Leaves.
|
|
|
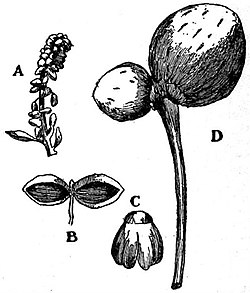
|
|
Fig. 12.—Ginkgo biloba. A, Male
flower; B, C, single stamens; D,
female flower.
|
|
Ginkgoales.—This class-designation has been recently proposed
to give emphasis to the isolated position of the genus Ginkgo
(Salisburia) among the Gymnosperms. Ginkgo biloba, the maidenhair
tree, has usually been placed by botanists in the Taxeae in the
neighbourhood of the yew (Taxus), but the proposal by Eichler in
1852 to institute a special family, the Salisburieae, indicated a
recognition of the existence of special characteristics which distinguish
the genus from other members of the Coniferae. The
discovery by the Japanese botanist Hirase of the development of
ciliated spermatozoids in the pollen-tube of Ginkgo, in place of the
non-motile male cells of typical conifers, served as a cogent argument
in favour of separating the genus from the Coniferales and placing it
in a class of its own. In 1712 Kaempfer published a drawing of a
Japanese tree, which he described under the name Ginkgo; this term
was adopted in 1771 by Linnaeus, who spoke of Kaempfer’s plant as
Ginkgo biloba. In 1797
Smith proposed to use the
name Salisburia adiantifolia
in preference to the “uncouth”
genus Ginkgo and
“incorrect” specific term
biloba. Both names are still
in common use. On account
of the resemblance of the
leaves to those of some
species of Adiantum, the
appellation maidenhair tree
has long been given to
Ginkgo biloba. Ginkgo is of
special interest on account
of its isolated position among
existing plants, its restricted
geographical distribution,
and its great antiquity (see
Palaeobotany: Mesozoic).
This solitary survivor of an
ancient stock is almost extinct,
but a few old and presumably
wild trees are recorded
by travellers in parts
of China. Ginkgo is common
as a sacred tree in the gardens
of temples in the Far East, and often cultivated in North America and
Europe. Ginkgo biloba, which may reach a height of over 30 metres,
forms a tree of pyramidal shape with a smooth grey bark. The leaves
(figs. 10 and 11) have a long, slender petiole terminating in a fan-shaped
lamina, which may be entire, divided by a median incision into
two wedge-shaped lobes, or subdivided into several narrow segments.
The venation is like that of many ferns, e.g. Adiantum; the lowest
vein in each half of the lamina follows a course parallel to the edge,
and gives off numerous branches, which fork repeatedly as they
spread in a palmate manner towards the leaf margin. The foliage-leaves
occur either scattered on long shoots of unlimited growth, or at
the apex of short shoots (spurs), which may eventually elongate into
long shoots.




