pericardium to the exterior, and these are the excretory organs or kidneys, formerly known as the organs of Bojanus. The walls of the pericardium are also excretory in parts, these parts forming the pericardial glands. In the majority of Mollusca the gonads are provided with a pair of ducts of their own. There are thus two pairs of coelomic ducts. This fact gives rise to the question whether the Mollusca are to be regarded as primitively segmented animals or not. In animals which exhibit typical segmentation or metamerism, such as segmented worms (Chaetopoda), each segment or metamere possesses its own coelomic cavity, a pair of coelomic ducts, and a pair of nephridia. The structure of the Mollusca in the greater number of cases agrees with the hypothesis that the primitive form was unsegmented, and therefore had but one pair of coelomic ducts and one pair of nephridia. In existing forms the latter disappear in the adult. In the most primitive forms of several classes there are no distinct genital ducts, the gonads when mature discharging into or through the kidneys. Among the Gastropoda, in the Aspidobranchia, there is no genital duct, and the gonad opens into the right kidney; in the more modified forms the left kidney alone is functional, the right has been converted into the genital duct. Among the Lamellibranchia again the kidneys serve as genital ducts in the Protobranchia and some Filibranchia. In the higher forms the opening of the gonad is shifted more and more towards the external aperture of each kidney until finally it is situated on the external surface, and thus the gonad secondarily acquires an independent aperture. In the Scaphopoda there is no distinct genital duct, the relations are as in Aspidobranchia. Among the Amphineura we find one pair of coelomic ducts in the Aplacophora, two pairs in the Chitons. In the former the genital coelom and the pericardial coelom are continuous and the reproductive cells escape by the renal ducts. In the Chitons or Polyplacophora, on the other hand, the two cavities are separate, and there are independent genital ducts. It is possible therefore to regard the latter condition as secondary, and to conclude that the separate genital ducts have been derived from the original single pair of coelomic ducts, as in Lamellibranchs.
The Cephalopoda, however, do not harmonize so well with this view. The earliest forms of this class geologically are the Nautiloidea. Assuming that these ancestral forms resembled the existing Nautilus in their internal anatomy, they had two pairs of renal ducts and one pair of genital ducts, which would apparently indicate, not a single metamere or unsegmented body, but three metameres. There are however only two pairs of branchiae. The Dibranchia, with only one pair of branchiae, one pair of renal organs, and one pair of genital ducts, are much more recent, not appearing till the end of the Secondary epoch, and therefore must be regarded as descended from the Tetrabranchia. The latter are represented in the Upper Cambrian formations, together with Lamellibranchia and Gastropoda, and there are no earlier Molluscan fossils than these. Palaeontology therefore throws no light on the question whether the metameric or the unsegmented Mollusca were the earlier. The development of the Cephalopoda affords at present no better evidence that the metamerism is secondary. That of Nautilus, which would be most important in this inquiry, is unfortunately still unknown. In the Dibranchia true nephridia have not been detected in the embryo, nor has it been shown that the genital ducts are derived from the renal tubes. On the other hand, there is no evidence that the forms which show no metamerism, such as the Gastropoda, are descended from metameric ancestors. On the whole, then, the most probable conclusion is that the original ancestral form of the Mollusca was unsegmented, possessed one pair of true nephridia, and one pair of coelomic ducts whose function was to conduct the generative products to the exterior. The chief types of Mollusca were already differentiated at the beginning of the geological record, and the metamerism which occurs in the Cephalopoda has been evolved within the limits of that class.
 |
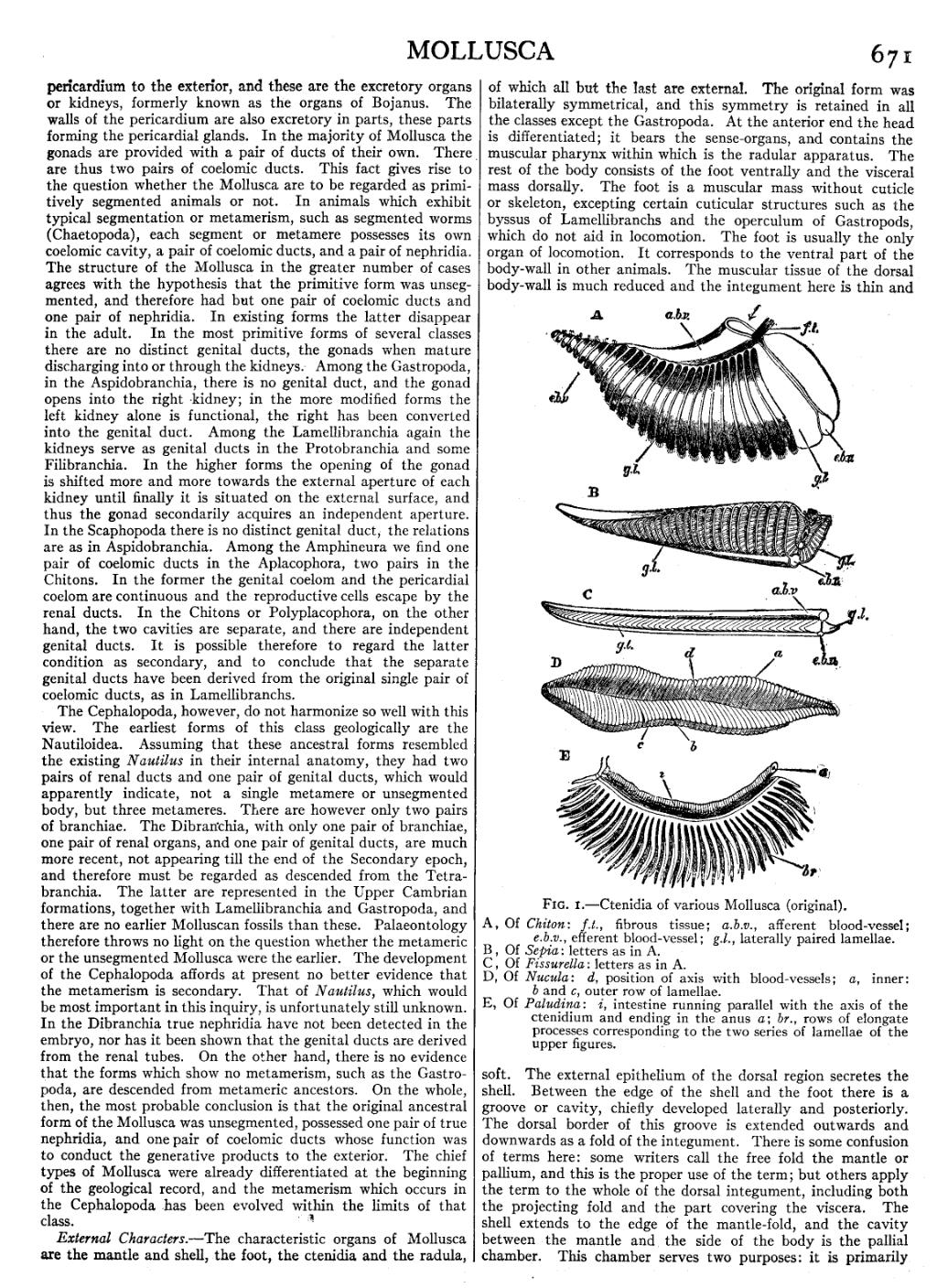
| Fig. 1.—Ctenidia of various Mollusca (original). |
|
A, Of Chiton: f.t., fibrous tissue; a.b.v., afferent blood-vessel;
e.b.v., efferent blood-vessel; g.l., laterally paired lamellae.
B, Of Sepia: letters as in A. C, Of Fissurella: letters as in A. D, Of Nucula: d, position of axis with blood-vessels; a, inner:
b and c, outer row of lamellae.
E, Of Paludina: i, intestine running parallel with the axis of the ctenidium and ending in the anus a; br., rows of elongate processes corresponding to the two series of lamellae of the upper figures.
|
External Characters.—The characteristic organs of Mollusca are the mantle and shell, the foot, the ctenidia and the radula, of which all but the last are external. The original form was bilaterally symmetrical, and this symmetry is retained in all the classes except the Gastropoda. At the anterior end the head is differentiated; it bears the sense-organs, and contains the muscular pharynx within which is the radular apparatus. The rest of the body consists of the foot ventrally and the visceral mass dorsally. The foot is a muscular mass without cuticle or skeleton, excepting certain cuticular structures such as the byssus of Lamellibranchs and the operculum of Gastropods, which do not aid in locomotion. The foot is usually the only organ of locomotion. It corresponds to the ventral part of the body-wall in other animals. The muscular tissue of the dorsal body-wall is much reduced and the integument here is thin and soft. The external epithelium of the dorsal region secretes the shell. Between the edge of the shell and the foot there is a groove or cavity, chiefly developed laterally and posteriorly. The dorsal border of this groove is extended outwards and downwards as a fold of the integument. There is some confusion of terms here: some writers call the free fold the mantle or pallium, and this is the proper use of the term; but others apply the term to the whole of the dorsal integument, including both the projecting fold and the part covering the viscera. The shell extends to the edge of the mantle-fold, and the cavity between the mantle and the side of the body is the pallial chamber. This chamber serves two purposes: it is primarily