
Creation by Evolution/Vestigial Organs
VESTIGIAL ORGANS
By George Howard Parker
Professor of Zoölogy, Harvard University
The body of an animal, like a piece of machinery, is made up of numerous parts that work together toward a common end. In an animal these parts are the organs, and each organ commonly has a definite use. Thus in man the eye, the hand, and the heart are three organs, the first for vision, the second for prehension, and the third for the propulsion of the blood. As in the machine, so in the animal, some parts are more important than others.
Just as there may be superfluous wheels and belts in a machine, so there may be organs in an animal that are not essential. A man may lose an arm or a leg and still live. Paired organs may be reduced by the removal of one. Thus a surgeon does not hesitate to excise a kidney provided its mate can be left intact. After such an operation the organ left behind usually enlarges and acts for two. Some single organs, such as the spleen, or even the stomach, may be removed without causing death. After its removal the function it ordinarily performs is taken over by other organs, and the life of the individual continues.
But in addition to organs of the kind just described there are others whose loss is followed by death. None of the higher animals can survive the loss of the heart, and every such animal requires at least the most of one lung and of one kidney. Even such inconspicuous organs as the adrenal glands, small bodies lodged in the fat near the kidneys, cannot be removed without causing death. Organs that are thus absolutely essential to the continuance of life are called vital organs and stand in strong contrast to others whose functions are not so essential. Different organs thus vary
-
 A
A -
 B
B
_p035_A_female_ant_with_normal_wings.png)
_p035_B_female_ant_with_vestigial_wings.png)
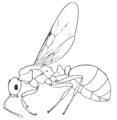
Fig. 1.—A, Female of an ant (Monomorium rothsteini) with normal wings; B, Female of an ant (Monomorium subapterum) with vestigial wings.
greatly in that some are essential and others are not essential to life.
Vestigial organs are those that are quite useless. Such organs are of course not necessary to their possessors. It is not always easy to prove that a given organ is vestigial. As might be inferred from what has been said, such proof requires that a given organ be not only removable without detriment from the animal of which it is a part but that it be shown to be without function. Conclusive evidence on this point is not always easy to obtain, for our ignorance may at times leave us in doubt as to the presence or absence of function in a given part. Notwithstanding this difficulty we know that many organs are truly vestigial, and such organs may be found in animals of almost any group.
Most insects are capable of flight and possess to this end a pair or, more commonly, two pairs of wings. Nevertheless many insects have wings that are entirely useless. Thus the male of the gipsy moth has well-developed wings and flies as other moths do; but the female, though she has fully formed wings, makes no use of them. When she emerges from her cocoon she creeps a short distance away and deposits
-
 A
A -
 B
B
_p036_A_claws_in_a_python.png)
_p036_B_skeleton_supporting_claws_in_a_python.png)
Fig. 2.—A, Claws at the sides of the vent of a python, representing vestigial hind legs; B, Skeleton supporting the claws. After Romanes.
her eggs, but without flight. Her wings are functionless and in that sense vestigial. In many other insects the wings are not only useless but are relatively small. Examples are seen in certain chalcids, small, almost microscopic wasp-like creatures that are often parasitic in other insects. Wheeler has recently described an Australian ant, Monomorium subapterum (Fig. 1), in which the wings of the female are about half the size of normal wings and are quite without function. Such wings are clearly vestigial.
In many cave animals the eyes are vestigial. Thus in a number of cave fishes the eyeballs are small, spherical bodies hidden under the skin and are of no use whatever. The eyes of certain subterranean crayfishes, insects, and salamanders are similarly useless and hence vestigial.
_p037_reproductive_organs_of_a_female_pigeon.png)
Snakes are commonly regarded as legless reptiles. But in the python (Fig. 2) a small claw can be see on each side of the vent, and these claws are supported by bones within the body in such a way that they are clearly vestiges of hind legs. Snakes are also peculiar in the structure of their lungs. Most air-inhabiting vertebrates have two lungs, one right and the other left. Many snakes have only the right lung, the left being represented only by a small protuberance. Vestigial organs are also well exemplified in birds. The ovaries and oviducts of most animals are evenly placed and equally developed on the two sides of the body, but in the birds these parts are functional only on the left side (Fig. 3). The ovary and the oviduct of the right side are abortive and quite useless, and hence vestigial.
The wings of certain birds are also vestigial. Wings used as organs of flight are among the most striking possessions of birds. Yet in the running birds, such as the ostrich and the cassowary, the wings are entirely useless for flight, for these birds are unable to rise off the ground. Relatively the feathers, bones, and muscles of the wings of running birds are very slight compared with those of the flying birds. In the New Zealand kiwi, the so-called apteryx, the wing is a diminutive member that is completely hidden under the feathers of the body, and hence the bird appears to be wingless. The wing of the kiwi is absolutely useless and is an excellent example of a vestigial organ.
Turning to the mammals we find many good examples of vestigial parts. Whales are mammals whose organization adapts them to a life in the sea. They are warm-blooded_p038_skeleton_of_a_whale.png)
_p039_homology_between_human_hand_and_horse_forelimb.png)
In the leg of the horse the vestigial splint bones are represented in solid black, and in the hand of man the corresponding bones are also shown in black.
The front and hind legs of the horse also contain vestigial parts. If the skeleton of the front leg of a horse is examined it will be found to be composed of many bones like those in the human arm. Buried in the flesh below the shoulder of the horse is a single bone corresponding to the one in our upper arm. Following this in the horse is a pair of bones duplicating the two bones of our forearm. Then comes, in both the horse and man, a small group of wrist bones. These are located in the horse at what is improperly called its knee. From the so-called knee in the horse to its hoof is a row of four stout bones, which correspond to the four bones in line with the middle finger of man. The first of these bones in the horse—that is, the one next the wrist bones—is the longest and corresponds to the long bone in the human hand that extends from the wrist through the region of the palm to the base of the middle finger. If this single long bone in the horse is examined it will be found to.have on either side of it delicate splint bones, which lie in the flesh of this portion of the horse’s legs but which support no parts in particular (Fig. 5). In position these two splint bones correspond to the long bones of our second and fourth digits, but they are not continued as such in the foot of the horse. The splint bones of the horse’s foot are obviously remnants of other digits and are in the strictest sense of the word vestigial organs.
Turning to man we find him no exception to the rule that vestigial organs are abundantly present in the organization of animals. The external ear of the human being is a complicated fold of skin supported within by cartilage or gristle and occupying a fixed position on the side of the head. So far as hearing is concerned it is probably an organ of no great value. At least its occasional loss works no serious detriment to the hearing of its owner. But in listening for very faint noises we commonly extend the ear by holding a hand behind it, so that this organ probably serves somewhat as a collector of sound.
Notwithstanding the functional insignificance of the human external ear, this organ is provided with a rather remarkable group of muscles (Fig. 6). Extending from what may be called its root outward to the surface of the head are three considerable muscles, whose respective actions would be to bend the ear forward, upward, and backward. In addition to these muscles the surface of the ear proper has upon it six or more small muscles whose contractions would change slightly the form of the ear. All these muscles are well developed in the ears of certain lower animals, such as the horse and the dog, in which the three extrinsic muscles direct the ear as an ear-trumpet might be turned in relation to a source of sound, and the muscles of the ear proper change slightly the shape of that organ to adapt it better, perhaps, for the reception of a given sound. The human ear is incapable of these movements. The muscles, though present, are ordinarily functionless, though occasionally a person will be found who can move his ears slightly and in this way demonstrate a limited control over some of these muscles, but even the movement he produces is so slight as to have no advantage whatever for hearing and to be rather a lusus naturae than an_p041_vestigial_muscles_of_human_ear.png)
Not only does the ear of man exhibit vestigial organs but a similar organ is found in the eye. Deep-seated in the nasal angle of the eye of man is a crescentic ridge of whitish tissue which, in consequence of its shape, is called the plica semilunaris or semilunar fold. It is not an organ that plays an important part in the action of the eye; in fact, it appears to be little more than a mechanical duplicature in the membrane in adjustment to the surface which it covers, and no one would suspect its meaning until he had examined the eyes of lower animals.
If we examine the angle of a cat's eye corresponding to the nasal angle in man we shall see there not a small fold of membrane but a veritable third eyelid. This additional lid may be made to pass under the two outer lids and over the whole exposed surface of the eyeball in a way to sweep this surface completely. It is provided with muscles by which this movement is quickly and easily made. This nictitating membrane, as it is called, is well developed in most mammals and serves to protect the eye. In man its representative is the insignificant fold already described, which, devoid of muscles and other appurtenances, is in its present condition a purely useless part, a vestigial organ. It is a noteworthy fact that, though the nictitating membrane is a functional organ in many mammals, it is reduced to a semilunar fold not only in man but in the monkeys and the apes.
Vestigial organs in man are found not only on the exterior of the body, as shown by the eye and the ear, but in its interior. In human beings the small intestine is not continuous with the large one end to end, but the small intestine enters the side of the large intestine, the natural termination of which is a pocket that projects backward and is known as the caecum. This pocket carries on its surface a small worm-like attachment, the so-called vermiform appendix, whose cavity opens directly into that of the caecum. In a way the vermiform appendix marks the real ending of the large intestine (Fig. 7). As is well known, the appendix readily becomes a center of intestinal disturbance and is the seat of the disease known as appendicitis, the usual surgical cure for which is to remove the appendix. Thousands of human beings have had the appendix removed and continue to live without experiencing any inconvenience. In fact, their position in life after the removal of this troublesome organ is so much more secure than it was before that it is now customary, as an incidental step in many abdominal operations, to remove the appendix on the assumption that a person is better off without it than with it. There is no reason to suppose that the appendix in man performs any function whatever, and since its removal is unattended by any subsequent inconvenience, it is commonly set down as a useless organ.
_p043_caecum_and_appendix_of_rabbit_and_human.png)
In other mammals than man the vermiform appendix or its equivalent presents a variety of conditions. It is not always easy in these lower forms to distinguish the exact limits between caecum and appendix, but in the rabbit, for instance, a large and highly complex caecum communicates freely and easily with an extended and apparently highly functional appendix. Without doubt these two parts are of great value in the digestive functions of this mammal, and the same may be said of them in many other animals. They are both greatly reduced, however, in the monkeys and in the anthropoid apes, where they are represented by a condition almost exactly like that in human beings. In consequence of the state of the vermiform appendix as seen in man it may be set down in the human species as a truly vestigial organ.
The so-called wisdom teeth of man under certain conditions partake of the nature of vestigial organs. As is well known, the milk dentition of a child contains in each half of each jaw two incisor or cutting teeth, one canine or eye tooth, and two molars or grinding teeth, making twenty milk teeth in all. When these teeth are shed a permanent tooth takes the place of each milk tooth and in addition three extra teeth appear in each half-jaw. These are the permanent molars, and their presence increases the permanent teeth to thirty-two. The last of these permanent molars at the back of each jaw is known as the wisdom tooth. Ordinarily the wisdom teeth are cut when the person is between twenty and twenty-five years of age, and they get their name from the belief that at that age the person has arrived at years of discernment. Not a few fail to cut these teeth or, in fact, even to form them at all. In such persons the number of teeth is four short of the usual total. Teeth that fail to cut the gums are of course useless and, in fact, like the vermiform appendix, they may be worse than useless, for such imperfect teeth may at times form centers of disturbance that call for surgical treatment.
This occasional reduction in the permanent dentition of man is in a way foreshadowed by what is seen in the monkeys. The new-world monkeys have a permanent dentition composed of a total of thirty-six teeth; the old-world forms, including the gorilla, chimpanzee, and other anthropoid apes, have four fewer permanent teeth and agree in this respect with man. Man appears to be going one step farther and to be reducing his dentition by dropping out another group of four teeth, the wisdom teeth, a step which, if finally taken, would place his permanent dental outfit at twenty-eight instead of thirty-two teeth. In man wisdom teeth that fail to cut the gums are, strictly speaking, vestigial organs, for under such conditions they are absolutely useless.
Not only is man the possessor of numerous vestigial organs in his adult state, but he also exhibits organs of this kind in his early stages of growth. Just within each nostril of the human embryo, or even of the new-born babe, is a small pore on the median wall of the nasal chamber (Fig. 8)._p045_Jacobson_organ_in_human_embryo.png)
Another vestigial feature, prenatal in time of occurrence, is seen in the lanugo of the human embryo, the covering of fine woolly hair found on the skin of the unborn human infant. This hairy covering is ordinarily shed before birth, and the separate hairs may often be identified in the amniotic liquor in which the embryo is immersed. This hairy covering is like that of the foetus of most mammals. At no time can it be of functional importance to the human embryo, and as it is ordinarily lost before birth it must be regarded as a vestigial organ, purely embryonic in its history. Its occasional retention after birth gives rise to the hairy men and women of the museums and the side-shows.
Man, both in his embryonic and his adult state, possesses an abundance of vestigial organs. In fact, students of this subject who have tabulated these parts have attributed to the human being almost a hundred such organs, and though some of these may on further investigation prove not to be true examples of vestigial parts, most of them certainly fall into this class, so that man may be said to be rich in organs of this kind.
The brief survey that has just been made shows that vestigial organs are widely distributed throughout the animal kingdom, and that they may be abundantly present in a given species, such as man. Their evolutionary significance has long been a matter of comment. If animals were specially created why should there be included in their bodies parts that are quite useless and often in fact positively detrimental to them? Why, for instance, should man possess a system of functionless muscles for his external ear, a useless hairy covering before birth, and a worse than useless vermiform appendix? No advocate of the theory of special creation has ever been able to give a satisfactory answer to these questions. To those who believe in special creation the presence of vestigial organs has proved a stumbling block that they have never been able to avoid. In fact, the occurrence of organs of this type has always been an insuperable obstacle to the acceptance of this view of the production of organic species.
From an evolutionist’s standpoint, on the other hand, vestigial organs are precisely what should be expected. They are organs in process of disappearance. In the course of evolutionary change certain organs might naturally lose their usefulness and be replaced by others of a more appropriate type. As such organs gradually decay, so to speak, they should be expected to appear in much the way that vestigial organs do. Man possesses a system of functionless muscles connected with his external ear because these muscles were once useful to that organ in a pre-human ancestor who had occasion to move his ears as some modern animals still do. Man has a hairy covering before birth in consequence of his derivation from a stock of animals once fully covered with hair. His vermiform appendix is the remnant of an organ that was once a functional part of the digestive system of a remote ancestor.
Like other animals, man is not only a highly equipped and efficient organism with a most marvelous system of parts adapted to serve his bodily needs but he is also a repository of some of the most interesting and important relics of the past, relics whose significance can be truly understood only if they are viewed from the standpoint of the evolutionist. These relics are vestigial organs, and it is in this way and in this way only that such organs can be understood.
Organic evolution is not a principle that is open to direct and simple proof. Like the movement of the earth around the sun it can be demonstrated only indirectly. We do not even know that the earth is round by direct inspection. The shadow cast by the earth on the moon in an eclipse, the appearance of the ship as it rises over the horizon, and a number of other occurrences in nature are best explained on the assumption that the earth is round. The Copernican theory explains astronomic phenomena, it accounts adequately for all happenings in the skies. In a similar way organic nature, plants as well as animals, is full of happenings that call for some general explanation, and no principle meets this call so surely as evolution does. Among these natural occurrences nothing is so difficult to understand, except from the evolutionary standpoint, as vestigial organs. These organs are really signs of the past; they afford as indisputable a proof of the correctness of the evolutionary view as can reasonably be expected.
REFERENCES
- Darwin, C. The Origin of Species. 1859. Many subsequent editions.
- Geddes, P., and Thomson, J. A. Evolution. New York, 1911.
- Holmes, S. J. Life and Evolution. New York, 1926.
- Lull, R. S., Ferris, H. B. and others. The Evolution of Man. 1922.
- Parker, G. H. What Evolution Is. 1926.
- Plate, L. Die Abstammungslehre. Jena, 1925.
- Romanes, G. J. Darwin and After Darwin. Chicago, 1892–1897.
- Weismann, A. The Evolution Theory. London, 1904.
- Wiedersheim, R. The Structure of Man. 1895.
- Wilder, H. H. History of the Human Body. 1909.
“Darwin himself would have turned his back on that theory (Evolution) if one single fact could have been produced in favor of the hypothesis of immutability, special creation, or supernatural agency. No such fact was forthcoming in his time, nor has any such fact been brought to light since.”—Dorsey.
Many kinds of animals and plants exist to-day that show no records, and many kinds that do not exist to-day have left their records, in the rocks. If the rocks tell a true story, the story they tell is Evolution.—Editor.